Piirialad ja võõramaa mehed
 Kuna viimastel nädalatel on elavnenud arutelu meie “identiteedi tuumikosa”, st. meie päritolu üle ning kuna ses osas käibib mitmesuguseid oletusi ilmselgest mütoloogilisest mõtlemisest (E. V. Saks) kuni ilukirjanduslike spekulatsioonideni (L. Meri) ja kuna kommentaator Salvey seda eraldi palus, otsustasin siia postitada väikese ülevaate, mille tegin 2009. a. lõpus. Võimalikele vigadele või ebatäpsustele tähelepanu juhtimine on teretulnud.
Kuna viimastel nädalatel on elavnenud arutelu meie “identiteedi tuumikosa”, st. meie päritolu üle ning kuna ses osas käibib mitmesuguseid oletusi ilmselgest mütoloogilisest mõtlemisest (E. V. Saks) kuni ilukirjanduslike spekulatsioonideni (L. Meri) ja kuna kommentaator Salvey seda eraldi palus, otsustasin siia postitada väikese ülevaate, mille tegin 2009. a. lõpus. Võimalikele vigadele või ebatäpsustele tähelepanu juhtimine on teretulnud.
Allpool refereeritud rahvastikugeneetika-alased uurimused tegelevad piirialadega: esimeste põlluharijate ning küttide-korilaste vahelise piiriga ning edasi juba Eesti ning selle naabritega, mis moodustavad soome-ugri maailma läänepiiri. Valdavas osas uuringutest on analüüsitud nii isaliinis edasi antava Y-kromosoomi DNA kui emaliinis edasi antava mitokondrilise DNA (mtDNA) markereid. Ajalooline rahvastikugeneetika on viimase 20 aasta jooksul üle maailma ja Euroopa väga suurt tähelepanu pälvinud. Eestis tegutsevad uurijad K. Tambets, R. Villems jt. Tegu on ühega vähestest loodusteaduslikest meetoditest, mis võimaldavad meil heita pilku minevikku. Keeleteadus võimaldab küll näidata erinevate keelte arengut ja seoseid, kuid selle leiud on väga raskesti dateeritavad. Arheoloogia jällegi tegeleb materiaalse kultuuriga ning arheoloogilise kultuuri mõiste sidumisel etnilise päritoluga tuleb eriti kaugemas minevikus olla väga ettevaatlik või sellest sootuks loobuda.
Kui mtDNA on igal vaadeldaval juhul euroopapärane, siis Y-kromosoomi DNA osas eristuvad haplogrupid, mis viitavad meessoost migrantide saabumisele – soome-ugrilastega seostatav N3 ning (indo)euroopapärased I ja R1a.
2009. a. oktoobris ilmus ajakirjas „Science“ Bramanti jt. uuring, mis heidab valgust Euroopa kõige varasemale põllumajanduslikule ajaloole. Küsimus sellest, mil määral toimus põllumajanduse levik koos migratsiooni ja mil määral oskuste levikuga, on olnud vaidluste keskmes pikka aega. Uurimisgrupp võttis emaliinis päritava mitokondrilise DNA (mtDNA) proove 22 küttide ja korilaste kultuuri esindajate jäänustest ning võrdles seda 25 prooviga varastelt põllumeestelt. Proovid pärinevad ajavahemikust 13 400 – 2 300 eKr.
Võrreldes kumbagi gruppi tänapäevaste eurooplastega, ilmnes tänapäeva Euroopa-sisese varieeruvusega võrreldes järgmisse suurusjärku kuuluv erinevus. Otsene järglus küttide-korilaste ja tänapäeva eurooplaste vahel tuleks kõne alla väga spetsiifilistes demograafilistes tingimustes, kui efektiivne naissoost populatsioon oleks pidanud olema alla 3000. See ei ole küll kooskõlas arheoloogiliste andmetega, samas on niisugust „pudelikaela“ ka raske välistada. Samuti on raske kindlaks teha seost efektiivse naissoost populatsiooni ja tegeliku populatsiooni vahel.
Suurem osa tänapäeva eurooplastest kannab haplogruppi H, V, U, J või T (pärinevad haplogrupp R-ist) või I, W või X (pärinevad haplogrupp N-ist). Kuigi mõni alamgrupp, nagu näiteks U5, on üldiselt omane vaid Euroopale, leidub suuremat osa nimetatutest nii Aasias kui Põhja-Aafrikas. Uuringu tulemusel ilmnes, et 82% küttidest-korilastest kandis haplogruppi U, väga valdavalt U5. Tänapäeva eurooplaste puhul esineb seda alates 1-5 protsendist inimestel Vahemere rannikult kuni üle 40 protsendini Skandinaavia saamidel. Kuna U5 on väljaspool Euroopat haruldane, võib arvata, et U-grupid olid jää-aja järgses Euroopas alates ajast 21 000 a. eKr tavalised.
Samas varaste põllumeeste proovides ei leidunud U5 ja U4 kandjaid ning levinumad olid N1a, H jt. tüübid. Küttide-korilaste proovides ei leidunud jällegi N1a ja H tüüpe. Järelikult on alust arvata, et põllumajanduse levik Euroopasse toimus koos märkimisväärse migratsiooniga, mille käigus vähemalt alguses ei toimunud rahvastiku segunemist. Autorid leiavad, et tänapäevaste eurooplaste geneetilist mitmekesisust ei saa hästi seletada vaid varaste põllumeeste ja küttide-korilaste populatsioonide järk-järgulise segunemisega.
Saamide küsimuses vaatleme järgnevalt Tambetsi jt. uuringut aastast 2004. Erinevalt Bramanti jt. uuringust katab see nii mtDNA kui Y-kromosoomi (päritakse isaliinis) markereid. Võrreldi 445 saami ja 17 096 Euroopa ja Siberi mtDNA proove ning 127 saami ning 2 840 Euroopa ja Siberi Y-kromosoomi proove. Ühe valdavalt saamidele omase haplogrupi U5b1b alusel analüüsiti 31 populatsiooni.
Arheoloogiliste andmete põhjal levisid arvatavasti Ahrensburgi kultuuri järglased Atlandi ookeani rannikut pidi Põhja-Skandinaaviasse (Komsa ja Fosna-Hensbacka kultuurid), Soome ja Koola poolsaarele ning Swidry kultuuri järglased Karjala ja Soome kaudu Skandinaaviasse. Nagu allpool näha, toetab geneetika seda teooriat vähemalt emaliini kohta.
Saami mtDNA proovides ilmnevad valdavalt (89%) V ja U5b (ainuke Euroopas laialt levinud grupi U5 alamgrupp) haplogrupid, mis on keskmise sagedusega levinud Euroopas Ibeeria poolsaarest Uuraliteni, kuid praktiliselt puuduvad obiugrilaste, samojeedide ja teiste põlissiberlaste hulgas. Aasiaga seostatavad haplogrupid D5 ja Z esinevad saamidel vaid 5% juhtudest.
Olgugi et on oletatud, et haplogrupp V pärineb Ibeeria refuugiumist, esinevad saamide hulgas haplogrupp V variandid, mis on enam levinud Ida-Euroopas, viidates võimalusele, et see on Fennoskandiasse levinud Ida-Euroopa kaudu. Samas puuduvad saamide hulgas V Edela-Euroopa ja Põhja-Aafrika variandid, mida leidub ka Saksamaal. Valdavalt saamidele omaseks peetud U5b1b1 alamgrupp on samuti levinud peamiselt Ida-Euroopas, mis annab alust arvata, et see kujuneski välja Ida-Euroopas.

Kaart: Tambets jt. 2009
Y-kromosoomi haplogrupid on kõigi saamide hulgas enamvähem ühtlaselt jaotunud, mis toetab nende ühist isapoolset põlvnemist.
Peaaegu poolel Y-kromosoomi proovidest ilmneb N3, mis on üldlevinud soome-ugri (balti 30%-40%, volga-soome 20-50%) ja Siberi populatsioonides, kuid peaaegu puudub Lääne- ja Lõuna-Euroopas ning ilmneb vaid vähesel määral (4-8%) rootslaste ja norralaste juures. Samas saamide hulgas kas puuduvad või esinevad harva haplogrupid C ja Q, mis on iseloomulikud samojeedidele jt. Siberi rahvastele, mongolitele, burjaatidele, kalmõkkidele, kasahhidele ja usbekkidele ning N2, mida leidub ainult volga-soome rahvaste juures. Seetõttu ei ole alust oletada hiljutist Y-kromosoomi voolu Siberist saami geenitiiki. N3 ja R1a (kokku ca. 60%) jõudsid Fennoskandiasse ilmselt Ida-Euroopast.
Teisel poolel juhtudest esinevad haplogrupid I, R1a ja R1b, mis on Euroopas laialt levinud. I esinemissagedus on ca. 1/3 ning see on laialt levinud kogu Euroopas, kuid peaaegu puudub Aasias (sh. samojeedide, hantide ja manside hulgas) ning seda defineeriv mutatsioon tekkis tõenäoliselt Euroopas.

Kaart: Tambets jt. 2009
Uuring ei toeta hüpoteesi, et saamid pärinevad põhjapoolsete uurali rahvaste või muude siberi põlisrahvaste hulgast.
Lätlaste kohta ilmus 2006. a. Plissi, Tambetsi jt. töö, milles uuriti 299 inimese mtDNA-d üle Läti. Levinuimaks haplogrupiks (45%) osutus H, mis on seda kõikjal Euroopas va. saamide juures. Umbes neljandik kuulus U gruppi, kõige enam U4 ja U5 alamgruppidesse (40%), mis on märkimisväärselt kõrgem Ida- kui Lääne-Euroopas. Üldiselt iseloomustavad kõigi kolme balti rahva mtDNA-d samad markerid kui kogu Euroopas (lähedased germaani rahvastele jt.-le) ning aasiapärase M haplogrupi alamgruppide D5b ja Z1 osa praktiliselt puudub. Samal ajal esineb neid soomlaste, karjalaste ja saamide hulgas 2-5%. Ungarlaste mtDNA-d uurides selgus Nádasi jt. uuringust, et aasiapäraste haplogruppide osakaal on samuti 5%.
Autorid toovad välja, et varasemad uuringud on näidanud Y-kromosoomi haplogrupi N3 kõrget, umbes kolmandikulist osakaalu. Sama käib ka eestlaste (30-35%) ja volga-soome (20-50%) rahvaste kohta. Samal ajal juba poolakate hulgas on selle esinemissagedus vaid 2-5% (nagu eelpool nägime, on ka 48%-lise N3 osakaaluga saamide naabrite rootslaste-norralaste hulgas see vähem kui 8%). Ääremärkusena võib mainida, et Põhja-Läti baltistus arheoloogia andmetel alles keskajal.
Kasperavčiūte jt. uurisid 2004. a. leedukate mtDNA ja Y-kromosoomi markereid. Analüüsiti 180 leedu mtDNA proovi ja 196 Y-kromosoomi proovi üle Leedu. Leedus on arheoloogilisi tõendeid võimalikust soome-ugri mõjust väga kasinalt ning soome-ugri veekogude nimetusi ca. 30. MtDNA proovid osutusid sarnaselt eelmise uuringuga lähedasiks kogu Euroopaga (peamised euroopapärased haplogrupid moodustasid 97%). Y-kromosoomi N3 haplogrupi osakaal oli 36.7% ja R1a 44.9%.
Nii lätlased kui leedukad on seega geneetiliselt lähemal soomeugrilastele kui teistele indo-euroopa rahvastele. Lätlased ja leedukad on ainsad indo-euroopa rahvad, kelle kohta saab niimoodi väita.
Viimaks, Palo jt. seadsid 2009. a. eesmärgiks uurida geneetilisi erinevusi Edela- ning ülejäänud Soome vahel kontrollimaks varasemat hüpoteesi, mille kohaselt teatud pärilike haiguste avaldumine ja homosügootsus just Ida- ja Kirde-Soomes on nende piirkondade asustamine 15.-16. sajandil väikese hulga inimeste poolt (nn. founder effect).
Selleks uuriti Y-kromosoomi, mtDNA ning autosomaalseid markereid (kanduvad edasi mõlemas liinis) 1126 meessoost soomlasel. Y-kromosoomi uuringute andmete mitmekesisus piirkondade kaupa osutus 5-10 korda kõrgemaks kui mtDNA ja autosomaalsete markerite puhul. Selline suur vahe viitab just meessoost isikute migratsioonile, samal ajal kui naissoost populatsioon jäi kogu maal suhteliselt ühetaoliseks ning tüüpiliselt euroopalikuks sarnaselt teistele Läänemeremaadele.
Uuringu tulemusena ilmnes, et Edela- ja Lõuna-Soomes (väljaarvatud Turu piirkonnas) ning Rootsi piiri äärsel Lapimaal on oluliselt kõrgem (25-30%) rootsipärase Y-kromosoomi haplogrupi I esinemissagedus. Kontrolliti ka vene päritolule viitavaid markereid, kuid tulemus osutus negatiivseks.
Kui arheoloogia andmetel toimus Soome asustamine kohe pärast jää taandumist 8500 eKr ning järgnevalt võib oletada migratsioone seoses kammkeraamika ja nöörkeraamika kultuuridega 5500 ja 2500 eKr, siis uuringu andmetel leidub kogu Soomes soome-ugri populatsioonile viitavat Y-kromosoomi haplogruppi N3, kuid 79%-ni tõuseb selle osakaal vaid Ida-Soomes, samal ajal kui mtDNA haplogruppide jaotus on üle Soome ühtlane.
Artikli autorid ei pea võimalikuks mtDNA haplogruppide ühtlase jaotuse kujunemist üle Soome vaid 20 põlvkonna jooksul, mis lahutab kaasaega Ida-Soome asustamisest, mis lisab kaalu arvamusele, et Skandinaavia migrantide hulgas olid suures ülekaalus mehed. Heaks näiteks sobib rootsikeelne Larsmo piirkond, millel ainukesena on rootsipäraste Y-kromosoomi markerite osakaal üle 50%, kuid mtDNA on lähedane ülejäänud Soomele.


Vasakul: Palo jt. põhjal. Hall – a priori eeldatud varane asustus (langeb kokku ka nöörkeraamika levikuga). Punane tähistus –HG I 24-30%. Kollane tähistus – HG I üle 50%. Katkendjoon – umbkaudne Rootsi-Novgorodi piir 1323. Paremal: Soome ilmateenistuse andmetel (http://www.fmi.fi/saa/tilastot_72.html#7) keskpäevaste üle +5 ulatuvate temperatuuride summa aastas 1971-2000.
Kokkuvõttes siis võib väita, et geneetika avardab jätkuvalt meie arusaamu varasematest rahvastikuliikumistest ning et iseäranis torkab silma varem käsitlemata võimalus, et teatud rännetes osales ainult üks sugupool. Kas ja kuidas muutus sel juhul rahvastiku (etniline) identiteet, on praegu raske öelda.
Kirjandus
Bramanti, B. jt. Genetic Discontinuity Between Local Hunter-Gatherers and Central Europe’s First Farmers – “Science” 2.10.2009: 326/5949, lk. 137–140
Tambets, K. jt. The Western and Eastern Roots of the Saami—the Story of Genetic “Outliers” Told by Mitochondrial DNA and Y Chromosomes – „Am J Hum Genet.“ apr. 2004: 74(4), lk. 661–682
Nádasi, E. jt. Comparison of mtDNA haplogroups in Hungarians with four other European populations: A small incidence of descents with Asian origin – „Acta Biologica Hungarica“ 2.07.2007: 58/2
Pliss, L., Tambets, K. jt. Mitochondrial DNA Portrait of Latvians: Towards the Understanding of the Genetic Structure of Baltic-Speaking Populations – „Annals of Human Genetics“ mai 2006: 70/4, lk. 439-458
Kasperaviciute, D. jt. Y Chromosome and Mitochondrial DNA Variation in Lithuanians – „Annals of Human Genetics“ sept. 2004: 68/5, lk. 438-452
Palo, J. U. jt. Genetic markers and population history: Finland revisited – „European Journal of Human Genetics“ 2009: 17, lk. 1336–1346
Kunagi olen vergruppide dieedi raamatut lugenud. Autor väitis seal, kui mul nüüd õigesti meeles on, et näiteks 0- veregrupi omanikud on küttide-korilaste vereliini jätkajad nö ja peaks siis toituma ka umbes sama moodi. Ei tea, kas need haplogrupid ja veregrupid peaks ka omavahel mingeid seoseid omama…
salvey ütles:
Natuke on, aga kuna peamises vere rühmitamise süsteemis AB0 kasutatavad antigeenid on inimese ajaloo mõttes kaunis iidsed — hulgast massilistest migratsioonidest ja segunemissündmustest märksa vanemad —, on need seosed enamasti pehmed korrelatsioonid, mitte teravad üleminekud. Õpetlik tabel on saadaval Blood Book’is: Racial & Ethnic Distribution of ABO Blood Types. Huvitavateks tähelepanekuteks on näiteks B- ja AB-veregruppide peaaegu täielik puudumine Austraalia aborigeenide seas ning väga madal esinemissagedus Ameerika indiaanlaste seas (ja on võimalik, et Ameerika indiaanlastel nende antigeenide taga peituvaid geene enne Kolumbust ei olnudki), iseäralikult kõrge AB-veregrupi levik Jaapani ainude seas ning ainult 0-veregrupi esinemine Brasiilias ja Boliivias elavate bororote seas.
Mis on loomulikult jama (The Skeptic’s Dictionary kirjutab lähemalt), umbes samasugune nagu mõnede jaapanlaste uskumus, et veregrupp ja iseloom on seotud. Jama saab lähemalt lugeda veebilehelt GenoType Diet: The Hunter.
Ilusad kaardid koos A- ja B-antigeenide alleelide esinemissagedustega kohalike populatsioonide seas on saadaval Palomar College’i veebilehel inimeste mitmekesisusest: Modern Human Variation: Distribution of Blood Types.
mõte, mis seda loo ülal osas asuvat kaarti uurides pähe tuli, oli selline, et õppigu aga venekeelne elanikkond koolis eestikeel ilusasti selgeks ja ongi sama hästi kui eestlased valmis. Sest geeenides on vahe ju üpriski minimaalne.( Ma ei mõtle seda, et nad ei peaks oma juuri ja ajalugu tundma vms.)
Venemaaga on esiteks nii, et ka ainult Venemaa Euroopa-osa niimoodi ühe territooriumina käsitlemine on väga ebatäpne.
Teiseks toetavad fakti, et Põhja-Venemaa oli vähemalt 13.-14. sajandini valdavalt soomeugrilise asustusega, ka nii arheoloogia kui kirjalikud allikad. Vladimir-Suzdalimaa (kuhu hiljem tekkis ka Moskva) oli samuti endine soomeugri ala, kuid venestus mitu sajandit varem.
Ühesõnaga on suur osa venelasi slaavistunud soomeugrilased. Ma oleks üllatunud, kui endine Novgorodimaa eraldi ei oleks geneetiliselt väga sarnane Baltikumiga.
Kaart muide ei ole ses mõttes kõige parem, et värvierinevused ei peegelda mutatsioonide kaugust üksteisest. Seda tuleks vaadata avalõigus tsiteeritud lehekülgedelt.
Naistest.
Üleüldse kiputakse migratsioonide ulatust tihti üle hindama. Näiteks Suure rahvasterännu ajal moodustasid sissetungivad barbarid ka vaid 5-10% kohalikust rahvastikust. Väga võimalik, et mtDNA osas kujuneski tänases mõistes euroopalik foon välja pärast põllumajanduse levikuga kaasnevat migratsioonilainet ja on sellest ajast peale enam-vähem samaks jäänud.
Selle kohta pilt:
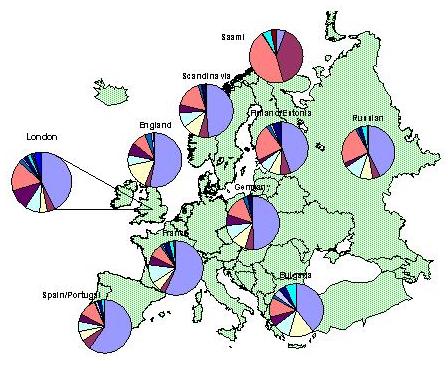
Maailma kohta:
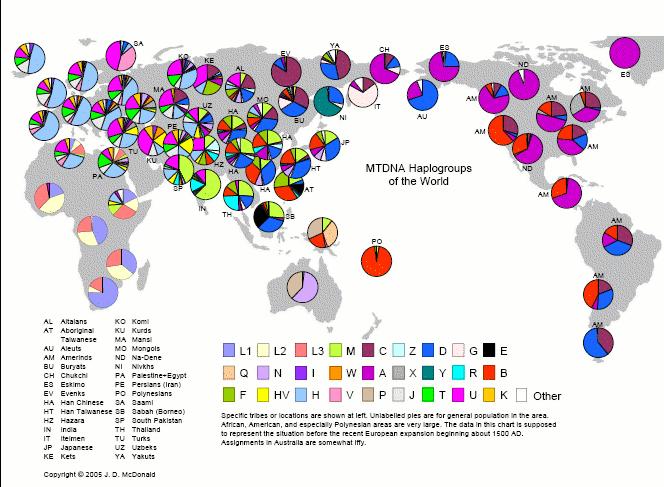
Allikas. Helesinine on siis mõlemal kaardil H.
Veel üks hea lehekülg: Mitochondrial DNA haplotypes for dummies.
Eesti on arheoloogia andmetel olnud püsivalt asustatud alates jääst vabanemisest ca. 9000 eKr. Tavaliselt soomeugrilusega seostatud kammkeraamika kultuur jõudis siia 4000 eKr. ning selgelt põllumajanduslik nöörkeraamika e. venekirveskultuur 3100 eKr (kuigi põllumajandus oli marginaalsel määral siia levinud juba ka enne). Kui ülejäänud Euroopale omane mtDNA jaotus tekkis Eestis ja Baltikumis seoses sellega, siis mida see ikkagi tähendab?
Kuidasmoodi muutus naisliini pidi edasiantav DNA üle Euroopa ühtlaseks, aga meeste hulgas jäi väga arvestatav varasem komponent püsima? Kas tõesti toodi valdav osa naistest pika aja jooksul põllumajanduspiirkondadest või siis nöörkeraamika asulatest (hilise kammkeraamika ja nöörkeraamika kultuurid eksisteerisid kõrvuti kiviaja lõpuni ning kammkeraamika kultuuri järglased õppisid muuhulgas ka omapäraseid kivikirveid tegema)? Mis siis enda omadega tehti? Ka siis, kui möönda, et U haplogrupi suhteliselt kõrge osakaal nii Eestis-Soomes kui Venemaal on jälg põllumajanduseelsest asustusest, on H ikkagi selgelt kõige levinum, siis tuleb tükk tühja maad ja siis kõik ülejäänud.
Kui aga oletada, et Baltikumi mtDNA sarnanes ülejäänud Euroopale juba enne N3 kandvate meeste saabumist, siis pidid seda levitavad migrandid põhja poole liikudes üle võtma U5 haplogruppi kandvate küttide-korilaste eluviisi ja nood siitkandist välja tõrjuma ammu enne, kui siin tegelikult põllumajandusega tegelema hakati.
Kui pärinemine põllumajanduslikust kultuurist andis pruutidele tõepoolest geneetilises materjalis nii tugevalt kajastuva eelise, kas etniline identiteet kandus siis edasi meesliinis ning vähemalt selle ajajärgu kontekstis peaks rääkima mitte emakeelest, vaid isakeelest?
Nagu näha, ei ole küsimustest meie “identiteedi tuumikosa” ümber puudust :) Vähemalt mina ei tunne aga küll oma identiteeti kuidagi ohustatuna.
Üks uuem artikkel (Haak jt. 2010), mis toetab Bramanti jt. järeldusi ning seob tänapäevase elanikkonna mtDNA markereid seni analüüsitud nöörkeraamika kultuuri matustest leitutega.
Pilt ka:
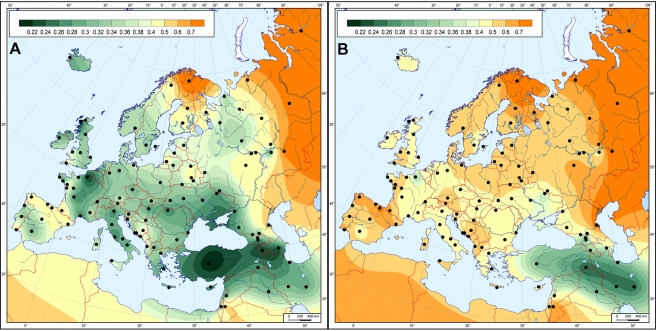
Vasakul siis tänapäeva Euroopa elanikkonna geneetiline kaugus kõigist nöörkeraamika kultuuri matuste seni analüüsitud proovidest, paremal tänapäeva Euroopa elanikkonna geneetiline kaugus nende poolt uuritud Derenburgi kalmistult saadud nöörkeraamika kultuuri matuste proovidest.
Nagu kergesti näha, on Eesti vasakpoolsel pildil kenasti skaala keskel ja kui Euroopa neljaks jaotada, ei tõuse kirdeveerandis kuidagi esile. Kindlasti mitte nii, nagu Y-kromosoomi N3 haplogrupp Baltikumis, Soomes ja Loode-Venemaal esile tõuseb.
Veel üks Venemaa Y-kromosoomi markerite kohta käiv artikkel (Balanovski jt. 2008).
Need kaardid on palju asjalikumad, kui artikli päises olev kaart.
(täna on siis päev mil minagi olen andnud oma tagasihoidliku panuse Google`i tõlke heaks, soovitades kaalikate asemel rootslasi ja turkside asemel türklasi. Ma loodan, et nad seda arvestavad, kuigi võimalik/ isegi tõenäoline (?), et keegi peale minu ei hakka seda lehte uurima Google`i tõlget kasutades.)
“Kuidasmoodi muutus naisliini pidi edasiantav DNA üle Euroopa ühtlaseks, aga meeste hulgas jäi väga arvestatav varasem komponent püsima?”
ma pakun hüpoteesina välja, et üleminekuga põllumajandusele kaasnes selline muutus mõtteviisis, mille tulemusena loobuti sündivuse piiramisest. ja kuna laste arv on siiski ennekõike naiste otsustada, võis tulemuseks olla põlluharijate geenide tugev domineerimine naisliinis.
ja selline väike märkus veel, et keel (ja kultuur) ei levi tegelikult geenidega. enamasti levib see prestiižiga. klassiühiskonna puhul (ja mõnikord võivad seisuste piirid joosta rahvusi või rasse pidi, näiteks indias on see nii olnud) võetakse üsna hõlpsasti üle valitseva klassi keel ja kultuur. prestiižiga levimise näiteks võib sobida ladina keele levik, või ka igasugused pidžinid ja kreooli keeled tänapäeval.
Pigem võiks arvata, et mitte ei loobutud sündivuse piiramisest, vaid palju rohkem vastsündinuid jäi ellu. Tõepoolest, neoliitilise revolutsiooni käigus kasvas Euroopa rahvaarv märkimisväärselt. Aga ses osas ei tohiks ju sugupooltel vahet olla?
Seda, kelle otsustada oli laste arv 5000 aastat tagasi, me ei tea.
Kindlasti ei levi keel ja kultuur tingimata geenidega. Nagu ülal osutatud, kiputakse migratsioone tihtipeale üle hindama. Etniline koosseis, arheoloogiline kultuur ning geneetiline taust on kolm ise asja. Need võivad olla lõdvalt seotud (Y kromosoomi haplogrupi N3 levik kipub tänapäeval soomeugrilaste asualadega kokku langema, mistõttu oletatakse retrospektiivselt sarnase seose olemasolu ka minevikus) aga ei pruugi (nagu tänapäeva lätlaste-leedukate puhul).
Keele koha pealt ei oska öelda, aga kui kultuur geenidest ei sõltu, siis millest ?
Kaaskommija “Starker” pakutud prestiiž ei kõla usutavalt. Euroopa mustlaste ning ja USA neegrite ajalugu näitab, et nad mitte ei võta prestiižikamat kultuuri omaks, vaid pigem vastanduvad sellele.
“Kultuur” meie igapäevatähenduses e. vaimne kultuur ja arheoloogiline kultuur on esiteks kaks ise asja. Arheoloogiline kultuur tähendab ühesuguse või sarnase materiaalse kultuuri levikut. Ses mõttes toimus Eestis järsk ja oluline nihe ka näiteks 1990. aastate alguses. Vaimset kultuuri saame jälgida siiski sellest hetkest alates, kui tekivad meetodid selle salvestamiseks (eelkõige kiri).
Teiseks, vaimne kultuur sõltub väga valdavalt kasvatusest ja keskkonnast. Võõra kultuuri kõrgema prestiizhi tõttu võidakse üle võtta neist emb-kumb või mõlemad. Materiaalse kultuuri puhul muidugi tihtipeale töötab ülevõetav kultuur paremini, aga ei pruugi. Eriti ei tasu teha kaugeleulatuvaid järeldusi arheoloogiliste kultuuride nimetuste põhjal. Nöörkeraamika pott polnud kammkeraamika potist millegi poolest parem ning nimetused pandi lihtsalt selle järgi, et keraamikat leidub alati kõige rohkem ja seda on kõige kergem määrata. Oluline vahe seisneb ikkagi selles, et esimene neist oli ilmselgelt põllumajanduslik.
Konspraator ütles:
Que?
ep ütles:
On Sul tõendeid selle kohta, et mõni mittepõllumajanduslik kultuur oleks “sündivust piiranud”? Minu meelest kõlab säärane hüpotees väga kahtlaselt.
@dig:
Mul on hüpotees, et kütid korilased ei kohtu nii tihti, kui karjakasvatajad. Jahimehed elavad palju hõredamalt, kui põllumehed. Kui mees on kaks nädalat jahil ja pere elab üksi keset metsa võib jääda mõnigi laps sigitama. Külas elavad mehed naised kogu aeg koos, suurem tõenäosus rasestumiseks.
Ehk kui kaks kultuuri asuvad kõrvuti kasvab põlluharijate arv veidi kiiremini.
Kammkeraamika kultuuri inimesed elasid ikka ka asulates. Need asulad koliti mõnekümne aasta pärast ümber (kui koht oli ära lagastatud). Metsa ja loomi oli võrreldamatult rohkem kui tänapäeval ja on kahtlane, kas kütid nädalateks kodust ära käisid. Rahvastiku kasvu pärast neoliitilist revolutsiooni seletab tõenäoliselt see, et rohkem lapsi jäi ellu. Ükskõik mis siin ka otsustav tegur ei olnud, ilmselt mõjutas see mõlemat sugupoolt ühepalju.
Kuidas aga seletada ülalkirjeldatud sugupoolte vahel ilmnevat geneetiliste markerite erinevust?
Kriku ütles:
Äkki kütid-korilased lõid põlluharijad-mehed lihtsalt maha ja omastasid siis nende naised?
Jah, see on ka üks võimalus nagu ka ülalmainitud põllupidajate hulgast pruutide toomine. Aga mis nad siis ikkagi oma naistega tegid? Saatsid Lapimaale?
dig ütles:
jah, seda on antropoloogid päris palju uurinud, näiteks inuittide ja ka mingitel muudel “primitiivsete” rahvaste juures. vastsündinute tapmist, eriti kaksikute ja tüdruklaste tapmist on peetud üsna sageli pruugitud sündivuse kontrolli all hoidmise meetodiks. (ma refereerin praegu peast paberilt loetud allikaid, osavate guugeldajate abi on teretulnud.)
ja see väide, et põlluharijatel jäi rohkem lapsi ellu. nojah, aga mis selle põhjus siis oli? arheoloogilised uurimised (skelettide analüüsid) viitavad enamasti pigem sellele, et põllumajandusele üleminekuga inimeste tervis üldiselt halvenes (toitainete puudulikkusest tingitud haiguste tõttu ja seetõttu, et rohkem rahvast koos tähendas rohkem nakkushaigusi) ja eluiga lühenes. mulle tundub siiski loogilisem mingi põhimõtteline muutus mõtteviisis, mistõttu sündivuse kontrolli ei peetud enam vajalikuks.
Vahva ülevaade ja arutelu.
Kui nüüd selle info kuidagi saaks Hans Roslingi Gapminderisse panna, vot siis oleks vast (populaarteaduslik) äge vaatamine! :)
(Pikem BBC saade Hans Roslingist)
minu “kõhutunne” ütleb, et see erinevus on kuidagi seotud asjaoluga et mitokondriaalne dna liigub ainult emaliini pidi (mitokondrid on munarakus, spermatosoidist läheb sügooti ainult tuum). St kui su emal oli “põllumajandaja” mtDNA, siis on ka sul, su vennal ja õel. Kui te oleta kultuuriliselt /isa kaudu/ “küttkorilased”, pärandab su õde järgmisse põlvkonda ikkagi põllumajandaja mtDNA. Ometi on tema ja ta tütarde puhul tegu “sinu hõimu” naistega, kes kultuuriliselt võivad ikka küttkorilased olla. Vennaga on keerulisem, tema mitokondrid on samasugused kui emal aga see mis on ta lastel, oleneb millise naise ta võtab.
Definitsiooni järgi saab see siiski olla vaid rahvastikukasvu mitte sündivuse kontrolli all hoidmise meetod, sest vastsündinud on ju juba sündinud. Aga muidugi ei saa välistada, et kammkeraamika kultuuri inimestel oli mingisugune norm oma tütarlapsi ära kustutada ja nöörkeraamika kultuuri omadel polnud.
See kõik on õige, aga see ei välista, et rohkem lapsi võis ellu jääda.
Minu teada kaasnes neoliitilise revolutsiooniga rahvastiku kasv igal pool – kas tõesti peaks eeldama, et igal pool oli enne seda kultuurinormiks rahvastikukasvu piiramine vastsündinute tapmise teel ja see norm kadus koos põlluharimise algusega ära?
Mis nakkushaigustesse puutub, siis need ongi ju tihedama asustuse, st. rahvastikukasvu tulemus.
Väike My, aga sarnane asi toimub ju ka meesliinis Y-kromosoomi puhul?
“kas tõesti peaks eeldama, et igal pool oli enne seda kultuurinormiks rahvastikukasvu piiramine vastsündinute tapmise teel ja see norm kadus koos põlluharimise algusega ära.”
no lihtsustatult võiks seda ju ka nii sõnastada. kuigi ma arvan, et rahvastikuprotsessid pole enamasti päris nii lineaarsed ja ühekomponendilised. ma pakuks pigem “paradigma muutust”. :D et kui kütid-korilased pidasid oluliseks oma rühma suuruse stabiilsena hoidmist (milleks tüdruklaste tapmine on üks ja ilmselt vähemalt mõnel pool kasutusel olnud meetod), siis põlluharijate ühiskondades seda vajalikuks ei peetud.
“See kõik on õige, aga see ei välista, et rohkem lapsi võis ellu jääda.”
aga mis selle põhjus siis oli?
Natuke samal teemal. Forte: kõik maailma keeled lähtuvad Aafrikast.
Kriku ütles:
Unarusse jätsid? :))
ep ütles:
Olen lugenud, et Hiinas visati tüdrukuid ära ka põlluharimisajajärgul, selle kohta peaks vist päris kindlaid andmeid olema küll.
Mart, aitäh huvitava artikli eest. Võrdlev foneetiline analüüs pole muidugi tema leiutis, vaid juba 20. sajandi alguses kenasti kasutusel.
Minul on jälle tõsiseid raskusi universaalsetesse kultuurinormidesse uskumisel.
Suurem stabiilsus, ennustatavus ja planeeritavus, mis mõjutas eriti imikute ja väikelaste suremust?
Viljelusmajandusel ju siiski olid märkimisväärsed eelised, kuigi keskmise inimese dieet võis kokkuvõttes kesisemaks jääda (mis on omakorda ka nii rahvastikukasvu kui suureneva ühiskondliku ebavõrdsuse tulemus). Viljelusmajandus ei toonud üleüldist paradiisi, aga suutis palju rohkem inimesi ära toita.
Kriku ütles:
ma juhiksin siinkohal tähelepanu sellele, et imikute tapmise tabu kui üldine kultuurinorm on suhteliselt hiline nähtus.
nii palju, kui mina põllumajanduse tekkimise kohta olen lugenud, on mulle jäänud mulje, et asi oli pigem vastupidi, et põllumajandusega kaasnes esialgu suurem ebastabiilsus, sest mõnest üksikust taimeliigist sõltumise tõttu suurenes ikalduse oht ja vähegi intensiivsema põlluharimisega käis kaasas ka muldade ärakurnamine ja erosioon (ehk siis algselt heal põllumaal hakkasid saagid langema). aga võibolla just sellepärast loobutigi sündivuse piiramisest, hakati tegema lapsi nö varuga. aga see ikkagi ei seleta põlluharijate naisliini laiemat levikut. kui lapsi jäi rohkem ellu, siis oleks ju loogiline arvata, et mõlemast soost lapsi jäi rohkem ellu.
ah jaa, ja varasemale jutule veel selline kommentaar, et tüdruklaste tapmine võib olla justnimelt sündvuse piiramise meetod. järgmises põlvkonnas nimelt. sest järglaste arvu määrab peamiselt viljakate naiste arv ja nende paljunemiskäitumine, meeste arvu mõju on oluliselt väiksem.
Seda küll, aga siin tuleb arvestada erinevate motiividega. Ma olen valmis uskuma, et vigaseid või väeteid või neid, keda nagunii ei oleks jõutud toita, tapeti igal pool, samal ajal kui näiteks soolise selektsiooni kohta tundub selline väide väga kahtlane.
Miks nad sinu arust siis üldse kõplama hakkasid? Oleks võinud ju jahti edasi pidada?
Jah.
Seda küll, aga ma olen skeptiline, kas mesoliitikumis niisugust social engineering‘ut kasutati.
Kriku ütles:
Sest tüüpiline (või keskmine või mediaan-) saak oli kõblates suurem kui odaga tarva järel silgates? See ei räägi kuidagi vastu põllumehe saagi suuremale ebakindlusele, mis tundub mulle täitsa mõistliku oletusena: põllult on võimalik ikka ümmargune null saada ja terve aasta on mokas.
“Miks nad sinu arust siis üldse kõplama hakkasid? Oleks võinud ju jahti edasi pidada?”
võibolla ei jäänud neil lihtsalt muud üle. minu mäletamist mööda tekkis põllumajandus kuivemapoolsetel rohumaadel ja võibolla seal oligi vähe neid taimeliike, mis inimesi toitsid. mistõttu tuli neid lihtsalt intensiivsemalt kasutada. ja rohumaade loomad kipuvad elama karjades ja olema väga ettevaatlikud.
“Ma olen valmis uskuma, et vigaseid või väeteid või neid, keda nagunii ei oleks jõutud toita, tapeti igal pool, samal ajal kui näiteks soolise selektsiooni kohta tundub selline väide väga kahtlane.”
aga miks? küllap käisid sinna juurde rituaalid ja mütoloogia, mille kohta on raske midagi teada saada. aga võibolla pole muinasjutud suurel hulgal neitseid õgivatest koletistest sündinud päris tühjale kohale.
aga millised võiksid olla alternatiivsed seletused? kui oletame, et küttide-korilaste juures mingi hulk naisi lihtsalt ei saanud lapsi, siis mis võis olla selle põhjuseks? mingid usulised tõekspidamised? matriarhaat? haigused? või teine võimalus, kui oletame, et põlluharijate seas oli mingil põhjusel suur hulk mehi paljunemisvõimaluseta, siis mis võis olla selle põhjuseks? haigused, mis vähendasid ainult meeste viljakust? sõjad või kahevõitlused meeste vahel, mille käigus mõlemad osapooled hukka said? rituaalsed hukkamised? matriarhaat? kõik alternatiivid, mida mina välja mõelda oskan, tunduvad mulle veel uskumatumad.
Põllumajandus tekkis igas maailmajaos eraldi (ja eri kultuuridega) ja levis kõikjale, kus seda praktiseerida andis. Ka siis kui tõesti mingis maanurgas ei oleks ilma kõplamiseta suudetud piisavalt toitu hankida, ei seleta see, miks Eestis kõplama hakati. Lisaks, kui mingis maanurgas olud äkitselt karmistuvad, ei lahenda seda põllumajanduse leiutamine – selleks ei jää enam aega – vaid väljaränne.
Minu meelest ei kannata mõte hüpoteetilisest universaalsest kultuurinormist teadaoleva paljususe taustal Occami Wilhelmi testi välja. Seda enam, et me otsime ju seletust puhtalt lokaalsele fenomenile.
Võimalik, et kunagi saame teada.
Arvata võib.
Siin on veel see nüanss, et põllumajandus käib alati koos koduloomade pidamisega, mis ka suurendab stabiilsust. Teiseks, seda, et kõik kultuurid korraga nii aia taha lähevad, et midagi võtta pole, tuleb siiski väga harva ette. Aga jah, eks siis oligi näljahäda ja surdigi massiliselt ära. 14. sajandi Euroopas toimunud kliimamuutus on hea näide.
Teisest küljest, nagu Amartya Sen on näidanud, hinnatakse ikalduse (tegeliku toidupuuduse) osa näljahädade puhul tihti üle.
ep ütles:
Seksuaalne valik? Võib-olla vastasid varajaste põllumeeste naised parajasti moes olevatele Willendorfi Veenusesse kätketud iluideaalidele mõnevõrra paremini kui küttivad ja korilevad naised?
See on võimalik küll, except that mõlemad ei pea ju ühe kakluse käigus surema. Täiesti piisav on, kui üks elab edasi, et järgmises kakluses surma saada.
Võimalik mehhanism: põllumajandusega kaasnenud suurem asustustihedus tõi kaasa suurema konfliktide esinemissageduse nii vara, seksi kui võimu taotlusel. Aja jooksul leiutasid inimesed ühiskonna, aga esimestel põllumajandajatel seda luksust veel ei olnud.
Hüpoteetiliselt võib ette kujutada ka võimalust, et mingitel põhjustel populariseerus inimeste kastreerimine.
See viimane nüüd mitte, sest see mõjutaks mees-, mitte naisliini.
Jah, naisliini mõjutaks ilmselt pigem see, kui mõni hõim asuks oma kivist nugadega naiste ümberlõikamist leiutama.
Kriku ütles:
*khm* monokultuur *khm*. Sotsiaalseid põhjuseid põllumajandusliku mitmekesisuse vähendamiseks on hulka, alustades vanast heast “naabri rukkisaak on suurem kui minu odrasaak” mõtlemisest ja lõpetades sellega, mis Iirimaal kartulinäljahäda kaasa tõi. Ja enne Kolumbust ju Euroopas kartuleid ei olnudki.
Kartuleid jah ei olnud, aga esimene Eesti õietolmuproovidesse ilmuv tänapäevane põllukultuur oli üleüldse kaer. On muidugi oletatud, et see kasvas nisu ja odra vahel umbrohuna ja eraldi seda veel ei kasvatatud. Lõuna pool läksid veel näiteks naeris ja põlduba söögiks veel enne põlluharimise algust. Kindlasti ei olnud Vahemeremaade põllumajandus neoliitikumis monokultuurne. Karta on, et suunda sinnapoole tekitasid arenevad ühiskondlikud suhted.
Ma ei oska muidugi öelda, kui suur oli just Eestis tõenäosus kõigi kasvatatavate kultuuride ikaldumiseks mingil hetkel viimase 4000 aasta jooksul. Usutavasti tõepoolest suurem kui rukki ja kartuli puhul. Ma ei ole seda kuskilt eraldi uurinud, millal just mis kultuurid Eestisse jõudsid. Kui mõni lugeja on, siis pangu kirja.
Nagu öeldud, käib põllumajandusega koos koduloomade pidamine. Kui ka mingi põllukultuur ikaldus, ei pruukinud hein ja kitsed ikalduda.
Kriku ütles:
Midagi ei teinud nad oma naistega. Kuna laste ellujäävus oli korilaste hulgas väike ja põlluharijate hulgas suur, siis suri korilaste liin lihtsalt välja.
See ka muidugi, et suguliitelistest geenidest on meieni jõudnud ainult need, mis on pärandunud katkematut samasoolist ahelat mööda. See lünklikkuse probleem sarnaneb mõnevõrra fossiilide kogumi ebatäiuslikkuse omaga.
Miks see antud juhul probleem on?
Sellepärast, et ta on võimalus geenialleelide kadumiseks ilma nende alleelide kandjate mittereprodutseerumiseta? Sellest, et mtDNA alleel X paar tuhat aastat tagasi geograafilises piirkonnas Y sagedusega Z levinud oli ja tänapäeval välja surnud on, ei saa tegelikult järeldada, et mtDNA alleeli X kandnud inimesed välja surid — ainult seda, et välja surid alleeli X kandnud mitokondrid. Seda võib põhjustada päris hulka põnevaid inimpopulatsioonilisi protsesse, millest kõige huvitavamad kestavad kümneid või sadu põlvkondi, mitte ei tähenda ühekordset kataklüsmi. Nukleaarse DNA Y-kromosoomi sisuga on põhimõtteliselt samamoodi, ainus erinevus on, et tollel ei ole eraldi ilusa nimega fenotüüpi.
Kriku ütles:
selle koduloomade pidamisega oli see jama, et neid ei peetud enamasti samas kohas, kus olid põllud. ja sõnniku kokkukorjamist ja põldudele kandmist eriti ei harrastatud. no polnud vist ka kuigi mõttekas tegevus, kui põllud olid all orus või jõe ääres ja karjamaad kõrgemal mägedes või lihtsalt mingitel kehvematel maadel kusagil eemal. sama probleem on mõnel pool aafrikas veel tänapäevalgi täiesti tõsine. sõnnikut kasutatakse mitmel pool pigem majade ehitamiseks või tule tegemiseks, eriti seal, kus puid on vähe. nii et see stabiilsuse suurenemine tuli vist alles mitmeväljasüsteemi kasutuselevõtuga. millal see oli? keskajal või uusaja alguses? igatahes ajaloolisel ajal, mitte eelajalooliste kultuuride juures.
ma hakkasin willendorfi veenuse peale mõtlema, et võib-olla oli põllumajanduslike ühiskondade naisliini edukuses oma osa viljakusriitustel ja sellega seoses naiste suuremal väärtustamisel. et kui küttidel-korilastel oli kombeks rasketel aegadel mütoloogilisi olendeid neitsitega sööta, siis põllumajanduslikes ühiskondades tekkis emajumala kultus ja koos sellega ka naise kui ema kultus. mis tõstis rasedate naiste ja emaduse ühiskondlikku prestiiži ja suurendas seeläbi laste arvu neis ühiskondades elavatel naistel.
Aga sellest ma ju räägingi…?
Igal juhul tõstab koduloomade pidamine stabiilsust seeläbi, et koduloomade näol on mingi toiduvaru olemas.
Kui ma nüüd Kriku argumendist õigesti aru saan, ei räägi ta mitte stabiilsusest väetamise kaudu vaid stabiilsusest tagavara-toiduallika kaudu. Tõepoolest, kitsed suudavad inimestele mittesöödavatest rohttaimedest kitsepiima ja kitseliha teha küll, heinaikaldus aga on madalama riskiga sündmus kui viljaikaldus.
ep ütles:
Keskajal.
Mitmeväljasüsteem muutus vajalikuks siis, kui pärast mulla ärakurnamist uue põllu teise kohta rajamine ebapraktiliseks muutus. Euroopas kasvas rahvastiku tihedus nii suureks esimeses lähenduses kaheksasada aastat tagasi. (Tegelikkus on muidugi keerulisem, kuna rahvas ei paiknenud homogeenselt ega kasvanud monotoonselt.) Mõned Aasia stepirahvad, kellel maad laialt käes, aga ei juurutanud mitmeväljasüsteemi enne 20. sajandit.
Mul on kuri kahtlus, et nendel hallidel aegadel ei olnud naiste ühiskondlik prestiiž teps mitte see pudelikael, millest rahvastiku juurdekasv otseselt sõltus.
Äkki põlduritel levid hoopis polügüünia? Ma nagu mäletan mingit uurimust Aafrika hõimu kohta kus see mees, kellel oli parem/suurem põld sai rohkem naisi.
dig ütles:
et kui neil leiba ei olnud, miks nad siis liha ei söönud? või küpsiseid?
see, et liha on peaaegu igaühel peaaegu iga päev laual, on siiski väga uus nähtus. ma olen aru saanud, et liha oli siiski peamiselt pidusöök ja ülikute söök. ilmselt sellepärast, et loomi peeti suhteliselt vähe. ja kuiva kliimaga aladel pole heinaikaldus madalama riskiga sündmus. kõrgmägedes võib olla, ja siin meie laiuskraadil ja meist põhja pool ehk ka, aga lähis-idas ja vahemere piirkonnas küll mitte. kui on põud, siis kõrbeb ikka kõik. kitsed ja kaamelid ehk saavad ka puukoort närides mõnda aega hakkama, aga lehmad on pirtsakamad. ja esimese asjana jääb toidunappuse korral piim kinni. kuigi lähis-idas pole piima tarvitamine vist kunagi eriti populaarne olnud, mul on kusagilt nagu meelde jäänud, et seal on laktoosi talumatus inimestel väga levinud. ja pealegi kui loomad raskel ajal maha tappa ja ära süüa, siis järgmine aasta neid lihtsalt pole, samas karja taastamiseks kulub pärast aastaid. aga kui põua-aastaid on mitu järjest?
Nagu öeldud, kui mitu aastat järjest kõik ikaldub, siis surebki väga palju inimesi maha. Aga seda tuleb siiski harva ette.
Nii Suurbritannias kui Maltal jätsid inimesed muide viljelusmajandusele üleminekul mereandide püüdmise järele. Ju siis said vajalikud proteiinid koduloomadest kätte.
See, kui palju neoliitilised inimesed liha sõid, on vaieldav. Viidatud artikkel on siiski oma järelduses selge:
Piimanduse kohta artikkel.
ep ütles:
Just. Leib sõltub konkreetsest kultivaarist või paarist, liha toormeks olev taimebaas on laiem ja ikalduse-risk madalam.
Ilmselt on see idealiseeritud mudel. Tõsi küll, ilmselgelt on liha osatähtsus (ja erinevate lihasortide osatähtsus) inimeste toidulaual ajatsi ja kultuuritsi varieerunud.
Enne põllumajanduse leiutamist oli ulukiliha oluline osa inimeste dieedist. Tõsi küll, kuna uluki kättesaamine ei pruukinud päris igapäevane olla, võib seda pidusöögiks pidada — kui jaht õnnestus, oligi pidu. Põllumajanduse juurutamine tõi esialgu kaasa enam-vähem püsiva toiduallika, aga kuna inimestel on statistiliselt paha komme oma riskikäitumist juhtida nii, et risk püsiks, järgnesid sellele arvatavasti tegevused, mis vanade toiduallikate mittekättesaamise-riski jälle tõstsid — näiteks jahikäikude harvenemine. Teatavate mööndustega võib rahvaarvu suurenemist ka riski ebapiisavuse kompenseerimise mehhanismiks lugeda. Keskaja kättejõudmisel oli juba aegu ja kohti, kus liha kallis ja haruldane, üsna palju. Aga keskajal oli ka hulka põnevaid epideemiaid, mis kippusid eelistatult ründama linnaelanikke, kes põllupidamisega vähem seotud olid ja sellepärast kukkusid keskajal põllumajandussaaduste hinnad aeg-ajalt madalale, kuna nõudlus langes ära.
Nagu klassikud ütlevad, määravad ajaloolisi protsesse aeg, ruum ja ühiskondlikud suhted (kui klassik seda juhuslikult lugema peaks, siis tervisi talle). Antud küsimuses on suur oht kanda varauusaegseid või keskaegseid ühiskondlikke suhteid, millest midagi kuulnud ollakse, üle kiviaega, millest me tegelikult mitte midagi ei tea. Arenenud ühiskondades nagu antiikmaailma impeeriumid, kõrgkeskaja ühiskond või hilisemad Euroopa riigid, otsustavad mõne toiduaine kättesaadavuse mitte looduslikud võimalused selle kasvatamiseks, vaid võimu- ja turusuhted. Näiteks liha osa eurooplase dieedis on palju kordi suurenenud-vähenenud (mul ei ole hetkel aega allikat üles otsida; kui keegi tahab, siis võin seda kunagi teha) ja nendes kõikumistes peegelduvad otseselt ühiskondlikud muutused. Ainuke looduslik põhjus, miks kiviaja inimene ei võinud iga päev liha süüa, tuleneb minu teada raskustest säilitamisel (Läänemere vesi on soolamiseks liiga mage).
Kriku ütles:
Kuivatamine?
Liha saab rasvaga ka sisse teha, aga AFAIK ei tulnud kiviaegsed eurooplased selle peale.